
Allozymsiche und chromosomale Ähnlichkeit bei zwei Drosophila-Arten
Maschinenübersetzung
Der Originalartikel ist in EN Sprache (Link zum Lesen) geschrieben.
Zusammenfassung
D. setosimentum und ochrobasis sind ein Paar sehr nah verwandter, teilweise sympatrischer Arten, die endemisch auf der Insel Hawaii sind. Männchen der beiden Arten unterscheiden sich auffällig im Flügelmuster, und es gibt Unterschiede in der Höhe und den Brutplätzen. Ähnlichkeitsindizes wurden für beide chromosomalen (C) und allozymischen (A) Varianten berechnet. Innerhalb der Hauptpopulationen jeder Art ergeben beide Datentypen Koeffizienten über 0,98. Der interspezifische Vergleich der Hauptpopulationen zeigt 0,66 (C) und 0,79 (A). Eine isolierte Population von ochrobasis aus den Kohala-Vulkanen (Ohu) zeigt im Vergleich zu setosimentum 0,68 (C) und 0,98 (A). Chromosomen sind somit viel empfindlicher als Allozyme, wenn es darum geht, diese Arten zu unterscheiden; das gleiche gilt für D. silvestris und heteroneura aus denselben Wäldern. Diese morphologisch unterschiedlichen Arten zeigen im Vergleich 0,96 (A). Alle vier Arten scheinen historisch gesehen sehr neu zu sein. In einem Gebiet sind etwa 2 % der wild gefangenen D. setosimentum/ochrobasis interspezifische Hybriden, obwohl ausreichende Proben darauf hindeuten, dass die separaten Genpools nicht zusammengebrochen sind. Die spezifischen Namen sollten beibehalten werden, aber die beiden Entitäten werden vielleicht am besten als recht fortgeschrittene Semispezies beschrieben, bei denen die reproduktive Isolation in der Natur jetzt nahezu vollständig ist.
Eine Revolution braut sich in der zeitgenössischen Systematik zusammen, insbesondere auf oder nahe der Ebene von Artunterschieden (1). Daten zur genetischen Variabilität innerhalb und zwischen eng verwandten Populationen können verwendet werden, um hochsensible Indizes genetischer Ähnlichkeit zu etablieren (2). Eine Übersicht über die biochemische Variation innerhalb und zwischen den Mitgliedern der Drosophila willistoni-Gruppe von Arten ist kürzlich erschienen. Eine direkte Beziehung zwischen Indizes genetischer Ähnlichkeit und bestimmten systematischen Bezeichnungen wurde vorgeschlagen (3).
Wenn Identität als 1,0 betrachtet wird, wurde gezeigt, dass lokale geografische Populationen einer Art durchschnittliche genetische Ähnlichkeiten von 0,97 aufweisen, Unterarten und Semispezies etwa 0,8, Geschwisterarten 0,52 und nicht verwandte Arten 0,35. Das vorliegende Papier berichtet über genetische Ähnlichkeiten innerhalb und zwischen einem Paar von teilweise sympatrischen, sehr eng verwandten Arten, die endemisch auf der Insel Hawaii sind. Wir führen die Verwendung von Indizes basierend auf chromosomaler Variabilität ein und vergleichen sie mit Indizes, die auf Alloenzymen basieren. Die biochemische Ähnlichkeit sowohl innerhalb als auch zwischen den Arten ist hoch, obwohl sie morphologisch und chromosomal stark differenziert sind. Diese Arten sind offensichtlich in einem historischen biologischen Sinne sehr neu. Wir schlagen vor, dass ihre biochemischen Ähnlichkeiten direkt mit diesem Umstand korreliert sind.
Materialien und Methoden
Proben von Drosophila setosimentum und ochrobasis wurden in feuchten Hochlandwäldern (1100-1600 m) auf der „Big Island“ von Hawaii gesammelt, wo sie endemisch sind. Sofern nicht unten ergänzt, wurden Details zu geografischen Lokalitäten, Sympatrie sowie zytologischen und elektroforetischen Techniken veröffentlicht (4). Neue Sammlungen wurden in Puu Makaala, 8 km nordöstlich von Olaa (Lokalität 11, Abb. 1 in Ref. 4) und an zwei Standorten entlang der westlichen Grenze zwischen dem Kau Forest Reserve und der Kahuku Ranch nahe dem südlichen Ende der Insel gemacht. Eines dieser Gebiete, genannt Kahuku Ranch - 3800 ft. (1150 m), lieferte D. setosimentum. Weitere neue Sammlungen, hauptsächlich D. ochrobasis, wurden in Kipuka Pahipa, 7,2 km nordöstlich des Kahuku Ranch - 3800 ft. (1150 m) Standorts gemacht. Dies ist dasselbe wie Lokalität 14, die in Ref. 4 beschrieben ist.
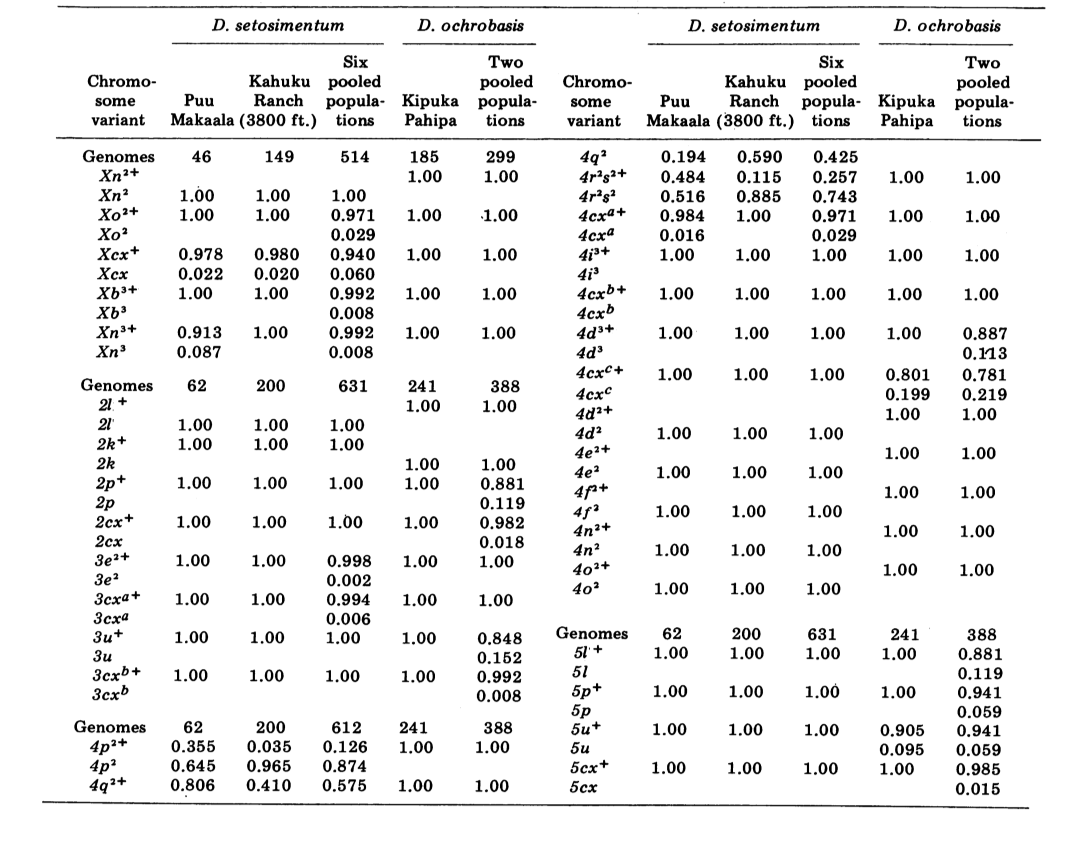
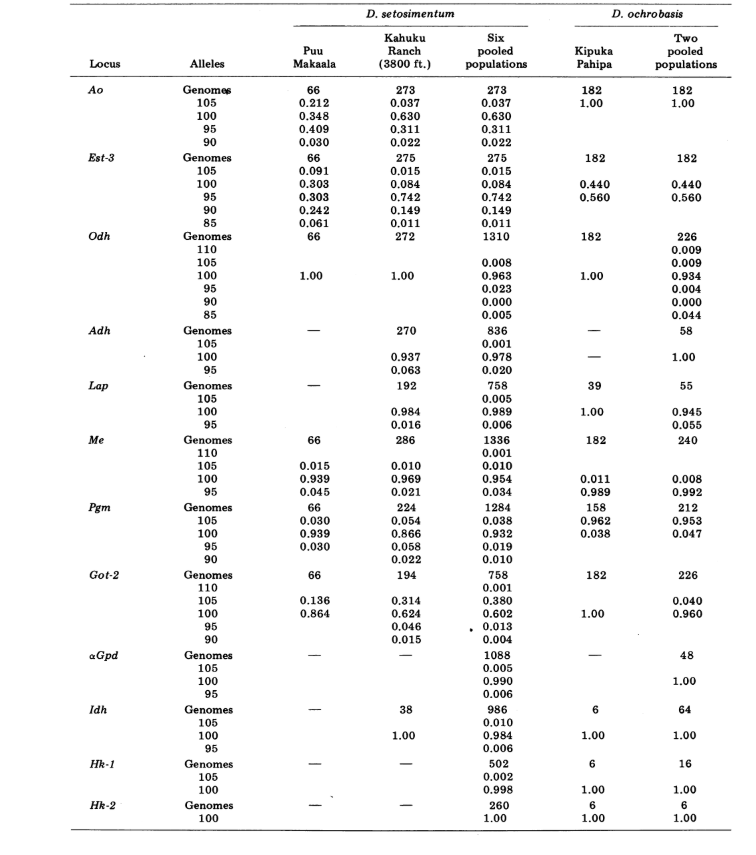
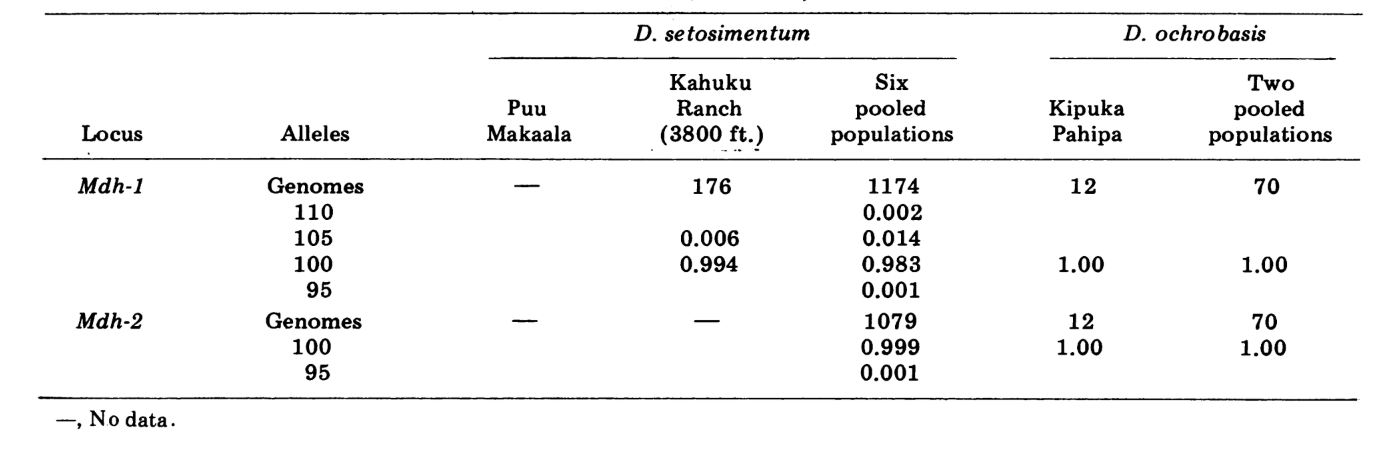
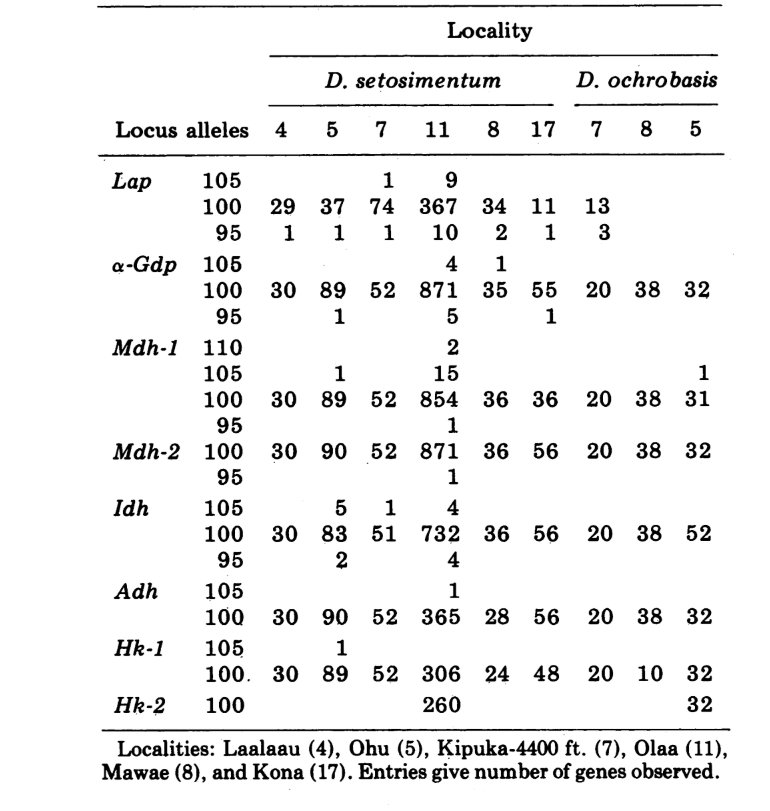
Wir berichten hier über Daten zu drei zusätzlichen Alloenzym-Loci, Aldehydoxidase (Ao), einem β-Naphthylacetat-Esterase-Locus (Est-3) und einem zusätzlichen Hexokinase (Hk-2), was insgesamt maximal vierzehn für den Vergleich von Populationen ergibt. Die beiden Arten zeigen Variation an insgesamt 30 polyten Chromosomenabschnitten aufgrund von Inversionen oder komplexen Chromosomen. Jeder Abschnitt wurde in jedem untersuchten Chromosomenabstrich der Speicheldrüse bewertet; das heißt, es wurde als heterozygot für die Variante, homozygot für die Variante oder homozygot für den Standardzustand aufgezeichnet. Diese Daten wurden verwendet, um Ähnlichkeitskoeffizienten auf die gleiche Weise wie für elektrophoretische Daten zu berechnen. Tabelle 2 listet eine neue Inversion, Xn3, von Puu Makaala. Eine erneute Untersuchung alter Folien von Mawae, Kipuka-4400 (1340 m) und Olaa zeigt, dass ihre Häufigkeit in diesen Populationen 0,056, 0,032 und 0,005 beträgt.
Beobachtungen
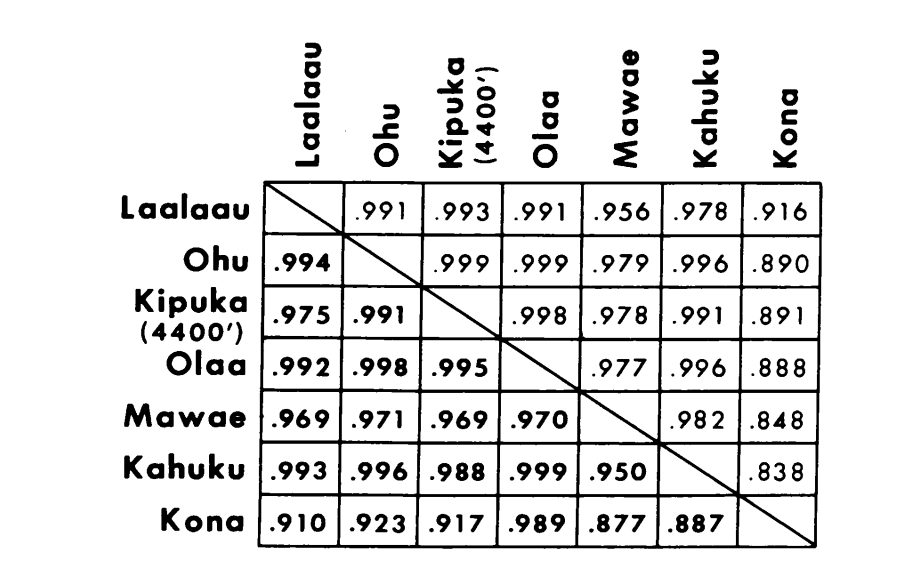
Die neuen Daten und die aus (4) wurden verwendet, um Ähnlichkeitskoeffizienten (Nei's I) für sowohl Allozyme (7-14 Loci pro Vergleich) als auch Chromosomen (30 Abschnitte pro Vergleich) zu berechnen, für sieben Populationen von D. setosimentum und drei von ochrobasis (Tabellen 1-3 und Abb. 1).
Die ersten sechs Populationen von D. setosimentum, die in Abb. 1 aufgeführt sind, stammen alle aus den Hochlagen der windzugewandten Seite der Insel. Sie zeigen einheitlich hohe Koeffizienten sowohl für Chromosomen als auch für Allozyme und können zusammengefasst werden. Der Mittelwert I mit Standardfehler ist in der obersten Zeile im Text von Tabelle 4 angegeben. Die meisten Vergleiche zeigen einige Unterschiede zur Kona-Population, insbesondere chromosomal (Abb. 1 und Tabelle 4). Letztere ist geografisch von den anderen auf der Südwestseite der Insel isoliert und weist einen festen Metaphase-Heterochromatin-Unterschied auf, der nicht in die Berechnung der polygenen Chromosomenkoeffizienten einbezogen ist.
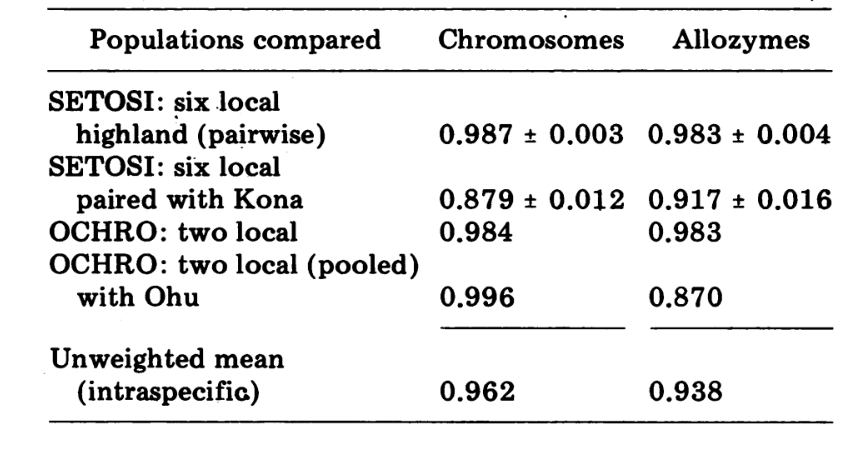
Die Daten von D. ochrobasis wurden auf ähnliche Weise behandelt (Tabellen 1-5). Die paarweise Untersuchung der beiden Populationen, Kipuka Pahipa und die kombinierten Mawae-Kipuka 4400 ft. Proben zeigen eine hohe Ähnlichkeit (Tabelle 4). Wenn diese Proben zusammengefasst werden, zeigen sie einen kleinen elektrophoretischen Unterschied zur ochrobasis Population von Ohu, die isoliert von den anderen am Kohala-Vulkan in der nordwestlichen Ecke der Insel ist. Die insgesamt ungewichteten intra-spezifischen Mittelwerte sind jedoch beide ziemlich hoch und ziemlich ähnlich (Tabelle 4).
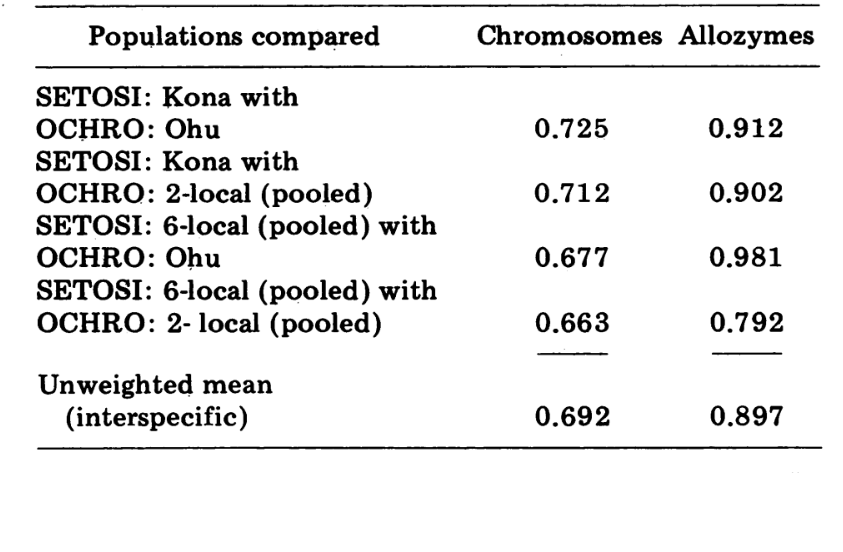
Inter-spezifische Vergleiche sind in Tabelle 5 dargestellt. Die Vergleiche in der Tabelle sind in der Reihenfolge abnehmender chromosomaler Ähnlichkeit aufgeführt. Die Daten zeigen, dass die inter-spezifischen chromosomalen Vergleiche viel effizienter darin sind, die Arten zu unterscheiden, als die auf Allozymen basierenden; tatsächlich liegen drei der vier letzteren Vergleiche über 0,9. Der Fall der Ohu-Population von ochrobasis ist interessant. Obwohl sie sich als unterschiedlich herausstellt, wenn Allozymen innerhalb ihrer eigenen Art verglichen werden (0,870, Tabelle 4), zeigt sie eine außergewöhnliche allozymische Ähnlichkeit zu setosimentum (0,981 mit dem Hochland-Pool und 0,912 mit Kona, Tabelle 5).
Diskussion
Obwohl die zytologische Differenzierung zwischen diesen beiden Arten erheblich ist, sind die biochemischen Unterschiede gering. Die beiden Entitäten scheinen jedoch keine Unterarten zu sein. Individuen jeder Art können in den meisten sympatrischen Situationen durch eine umfangreiche Reihe fester Inversionen klar erkannt werden, und es gibt auch Höhenunterschiede (4) und Unterschiede in den Brutgebieten (5). Männchen der beiden Arten unterscheiden sich auffällig im Farbmustern auf den Flügeln (6).
Andererseits gibt es erhebliche Beweise, die darauf hindeuten, dass D. setosimentum und ochrobasis als Semispezies betrachtet werden könnten (7). Die Schlussfolgerung, dass in der Vergangenheit eine gewisse Hybridisierung zwischen ihnen stattgefunden hat (4), wurde nun durch eine kürzliche Entdeckung (8) bestätigt. Im Gebiet des Kahuku Raneh-Kau Forest Reserve nahe dem südlichen Ende der Insel wurden ein natürlich vorkommender Fl und drei Backeross-Hybriden zwischen den Arten eindeutig erkannt. Dies macht etwa zwei Prozent der in diesem Gebiet untersuchten Wildfliegen aus. Angesichts der erheblichen natürlichen Proben, die untersucht wurden, und der Fülle sowohl an elektrophoretischen (9) als auch an zytologischen Markern in diesem Gebiet kann das Interbreiten als eher spärlich beurteilt werden.
Sekundäre Geschlechtsunterschiede, wie sie die Männchen dieser Arten kennzeichnen, sprechen nicht gegen den Vorschlag eines Semispezies-Status. Tatsächlich unterscheiden solche Merkmale einige der klassischen Fälle von Semispezies, wie zum Beispiel bei bestimmten Paradiesvögeln (7). Wie bei den Vögeln beziehen sich diese sekundären Geschlechtsmerkmale auf das komplexe territoriale (Lek) und sexuelle Verhalten, das von vielen der „Bildflügel“-Gruppe der hawaiianischen Drosophila, zu der diese Fliegen gehören, manifestiert wird (10).
Schließlich ergibt der umfassendste elektrophoretische Vergleich (4. Vergleich, Tabelle 5) einen Wert von 0.792, der nahe dem für die Semispezies von Drosophila paulistorum vorgeschlagenen Wert liegt (3). Im Gegensatz zu letzterem scheinen jedoch D. setosimentum und ochrobasis am besten als recht fortgeschrittene Semispezies beschrieben zu werden, bei denen die reproduktive Isolation in der Natur noch unvollständig ist. Wir befürworten die Beibehaltung der beiden spezifischen Namen für die beiden Entitäten.
Es bleibt jedoch die interessante Tatsache, dass bestimmte Populationen der beiden Arten elektrophoretisch praktisch nicht unterscheidbar sind. Die extremste davon ist die Ohu-Population von ochrobasis, die einen Ähnlichkeitsindex von 0.981 aufweist, wenn sie mit D. setosimentum verglichen wird. Die parallelen zytologischen Informationen zu dieser Population schließen die Möglichkeit aus, dass die elektrophoretische Ähnlichkeit auf aktuelle Hybridisierung zurückzuführen ist.
Eine enge interspezifische elektrophoretische Ähnlichkeit ist nicht unbekannt zwischen anderen Arten von Drosophila von der Insel Hawaii (11). Zum Beispiel weisen die sympatrischen Arten D. silvestris und D. heteroneura umfangreiche morphologische (12) und zytologische (13, 14) Unterschiede auf. Sie zeigen ethologische Isolation (15) sowie andere biologische Unterschiede (16). All dies impliziert einen erheblichen genetischen Unterschied zusätzlich zu den Inversionen. Dennoch zeigen diese beiden Arten einen allozymischen Ähnlichkeitskoeffizienten von 0,96 (11). Laut den Vorschlägen in (3) würde dies sie als lokale Populationen derselben Art einstufen, eine unhaltbare Interpretation.
Die Insel Hawaii ist geologisch neu (weniger als 7 x 105 Jahre). Arten, die endemisch auf der Insel sind, sind notwendigerweise noch neuer. Einer von uns (17) hat vorgeschlagen, dass die Artbildung in diesen und anderen Formen möglicherweise eine erzwungene genetische Umstrukturierung nach den stochastischen Effekten eines allopatrischen Gründerevents umfasst hat. Im Gegensatz zu den Themen dieses Papiers sind jedoch einige Fälle bekannt, in denen Arten ohne erkennbare polytene Chromosomenumstrukturierung gebildet werden. Ein Cluster von acht (jetzt 9) homosequentiellen Arten, die mit D. grimshawi verwandt sind, ist bekannt (18). In ähnlicher Weise zeigen die vorliegenden Fälle, dass Arten tatsächlich auch mit nur einer geringen Menge an allozymischer Umstrukturierung gebildet werden können. Wir schlagen vor, dass dies mit der Neuheit der artbildenden Ereignisse zusammenhängt. Möglicherweise sind die beiden Populationen gerade erst aus einer stark stochastischen Phase in ihrer Lebensgeschichte als Arten hervorgegangen. Diese Phase könnte hauptsächlich durch rekombinative Umstrukturierung im Verhältnis zur Stammart und die Etablierung neuer intern ausgewogener genetischer Systeme gekennzeichnet sein. Die Phase der neuen adaptiven Reaktion und der Umweltverfolgung könnte gerade erst begonnen haben.
Diese Sichtweise betont auch einen Punkt, der kürzlich in einer Diskussion über die auffälligen biochemischen Ähnlichkeiten zwischen Mensch und Schimpanse (19) angesprochen wurde. Speziationsereignisse können regulatorische Gene umfassen, die durch aktuelle Methoden der Populationsgenetik normalerweise nicht offenbart werden.
Autoren: H. L. Carson, W. E. Johnson, P. S. Nair, F. M. Sene